
All right for this post - and this is the one I hinted to a while back with allusions to imitation crab meat - I want to discuss a seldom mentioned idea for the neck. Now I don't know who first suggested this idea, I do know I first came across it on Adam Smith's post for "Why did elasmosaurids have such a long neck?" The suggestion, which at first notion might appear a little ludicrous, is that the hyper long neck of elasmosaurids (and maybe other plesiosaurs) could serve as a (Patreon) food storage device.
 |
| credit Adam Smith from plesiosauria.com |
 |
| Leatherback papillae. wiki commons |
 |
| Leatherback eating pyrosome credit Brian Skorm |
As I have stated before I think the biggest dietary signal we get regarding plesiosaur (long necked) is opportunistic - and with dietary items ranging from marine reptiles, pterosaurs, shellfish, scavenging, and crustaceans - that is a pretty defensible stance. Additionally I am of the bias that, on a whole, the predatory reptile brain is a lot less picky when it comes to diet than many derived mammal or even bird predators. Reptiles think but two thoughts; 1) Can I overpower it? and if so 2) Can I get it in my belly?
But what about choking? It's one thing for the leatherback sea turtle to stuff itself full of squishy stuff, but if plesiosaurs are eating shelled stuff, vertebrates, etc etc., can't that stuff block the windpipe?
True, but one solution would be for plesiosaurs to have an elastic glottis that connects to the trachea and just as snakes can breathe through it while swallowing large prey so could a plesiosaur snatch a breathe with a throat stuffed full of food. As an added cool point a pliable glottis would allow plesiosaurs to "hiss" as snakes do with theirs.
 |
| Glottis of boa swallowing. credit reptile-parrots.com |
Anyways, the point was to have fun with this idea. There is no reason I can think of to discount the neck as a food storage device and some arguments to be made for it. I guess it's up to you...
But for the second part of this post I want to discuss the "trap guild"plesiosaur characterized by having fine, homodont, interlocking toothed dentition, and provide an augmentation to the already established idea that this guild strained food from the water column/substrate. That is the neck itself played an active role in prey harvest beyond stealthy approach. What I am suggesting is that the oesophagus could, in conjunction with the gular region, expand to accept large volumes of water with prey. And then, shutting the mouth and closing the finely meshed teeth, expel this water back out through the shut mouth. The fine, needle like teeth of this morpho-ecological guild retaining any organisms that were taken in with the water.
 |
| Aristonectes lunge feeding on krill. Duane Nash |

I) Suitable prey is detected
II) Jaws are opened to receive prey and water. Powerful and rapid flipper thrusts propel the animal toward food patch. Engulfment of water forces open gular pouch and oesophagus.
III) Gular pouch and oesophagus further expand as the animal continues forceful swimming.
IV) Mouth is shut upon maximum volume achieved. Forceful swimming diminishes.
V) With mouth firmly shut contraction of oesophagus and gular pouch force water out while prey is retained behind finely, meshed teeth. Prey is swallowed after water is voided.
This scenario does invoke a classic question in paleo-marine reptile ecology. Why no baleen whale/giant filter feeding equivalents among marine reptiles? I have pondered this a lot. We now have evidence of a previously largely cryptic Mesozoic lineage of filter feeding fish. And ammonites might have taken up this niche. We will revisit this idea later...
Here I want to make an important distinction between two general strategies often times put under the banner of filter feeding. First of all is the more passive filter feeding - best exemplified by the giant Bowhead whale and basking shark. These animals, jaws agape, slowly cruise after and eat abundant microscopic organisms, copepods, etc, etc, which they are constantly filtering through their baleen and gill rakers respectively.
 |
| Basking Shark feeding. wiki. credit Conscious |
 |
| Humpback feeding on yoy pollock. Alaska. wiki. credit NOAA |
Let us explore this feeding method with the genus (and probably family) of derived elasmosaur, Aristonectes. These animals are very peculiar indeed and don't get the attention they deserve compared to other plesiosaurs and marine reptiles. My reasoning for this is that they are not big fanged and nasty looking. And that they come from the southern hemisphere while the vast majority of paleontologists /hobbyists/fans come from the northern hemisphere. There is that. And they are also relatively new to science. Let's do a quick primer on what we know.
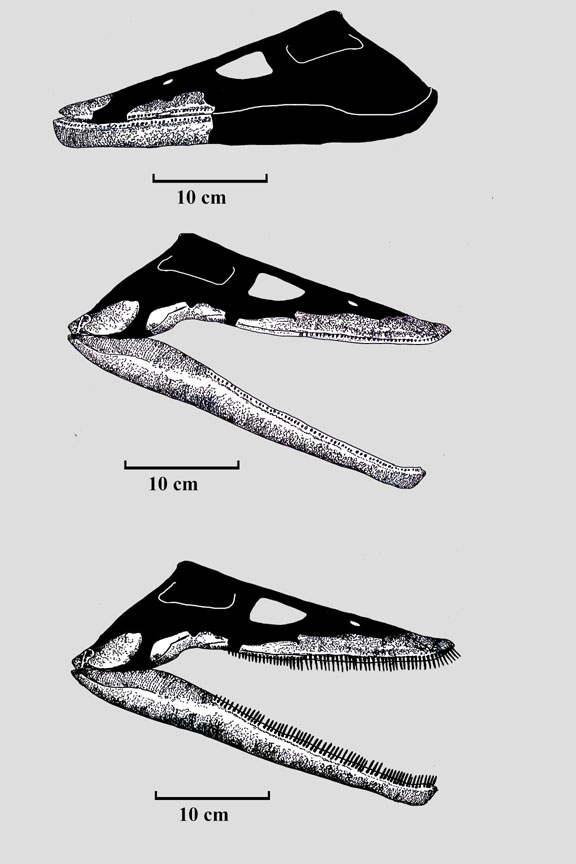

Same specimen as above but this ventral view really exposes the relative width of the jaw. If, as these animals are sometimes suggested to be, they were specialists on soft bodied squid why not grow the jaw longer to resist water resistance and expand reach when snapping shut? And I am not suggesting that they could not hunt squid - far from it I always assume opportunistic foraging unless shown compelling reasons otherwise - but I don't see how this design is optimized for that lifestyle.
 |
| Aristonectes. Chaterjee & Small 1989 |
 |
| Aristonectes. Chatterjee & Small 1989 |
Now, with these anachronisms in mind (small, pin like interlocking teeth combined with broad jaws) let us move on to the rest of the skeleton in these guys and see what other clues they might reveal.

Abstract for above diagram:

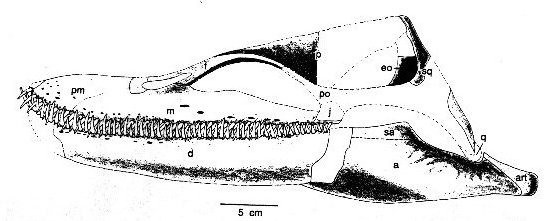
Ok, so the phylogenetic part is interesting and suggests Aristonectines are derived elasmosaurs but now onto the morphology: "It is a large plesiosaurian with a relatively large skull with numerous homodont teeth, a moderately long and laterally compressed neck, and relatively narrow trunk, with slender and elongate forelimbs."
Now again, some anachronisms with the head and neck. If you were a small game specialists of squid, small fish why evolve a relatively large skull but retain pins and needles for teeth? And then why a laterally compressed neck? The laterally compressed neck is inferred from the relatively diminished transverse processes of this particular plesiosaur - which also highlights the fact that most plesiosaur necks are not restored muscular/thick enough as well. Better reframed this way: Why diminish the lateral musculature of your neck if you are specialized for darting after small prey? Wouldn't you want ample musculature for capturing such a prey base? However if you are adapted primarily to swim straight into vast shoals of food a diminished lateral neck musculature is not a problem.
Also why, in the illustration above, do the transverse processes along the neck trend towards the anterior of the animal?

 |
| Crytoclidus. credit Adam Smith plesiosauria.com |
Note how in Cryptoclidus (below), another member of the "trap guild" although only distantly related, we see the transverse processes as basically horizontal to the vertebrae or trending posterior...? It's interesting and perhaps speaks towards some reorganization of the neck musculature in aristonectines perhaps due to its feeding specialization.
The name Aristonectes literally translates into "best swimmer" and there are some features suggestive of that description being apt. Note the description of above aristonectines having a relatively narrow trunk. Now, as I have discussed before, a lot of illustrations of plesiosaurs are too gaunt and feature the shrink-wrapped look, but here with aristonectines we see this group was relatively less round shaped than other plesiosaurs. In fact with this elongated look I am reminded of another clan of lunge feeders - rorquals - which are relatively sleek and streamlined (especially compared to passive filter feeders like Bowheads). The narrow trunk, slender and elongate forelimbs may have been adaptations in aristonectines, that allowed for efficient transit to and after highly dispersed but concentrated feeding patches.
 |
| Minke Whale Orange county coast CA. credit & (c) Shane Keena |

Again what I am describing is none too dissimilar from what may be described as general consensus that Aristonectes was more or less analogous to something like a crabeater seal - which uses specially derived cusped teeth to filter krill from its Antarctic waters. What is new in my interpretation and which has never been suggested before to my knowledge is that the gular region/oesophagus expanded to hold the water/food prior to being pushed back through the teeth.
 |
| Crabeater Seal. wiki |

A little thought experiment. What is the simplest, most parsimonious (fewest steps needed) way to turn a base model elasmosaur - imagine something like Thalassomedon - and turn it into an efficient filter feeders? It would prove very cumbersome and, in my opinion, outside the bounds of Darwinian evolution, to invoke the evolution of complex facial musculature, innervation, and so on to create a mode of filter feeding exactly the way mammals do it. Instead simply arranging the oesophagus and gular area into a food storage area - and co-opting the musculature already there to assist in pushing water back out through the sieving teeth is the simplest way to achieve this adaptation. No need to make the buccal area highly complex with large mobile tongue, muscles/nerves etc etc - simply through using what is already there to best fit - exaptation - you got yourself a pretty nice lunge feeding filter feeder equivalent to rorquals for all intents and purposes.
 |
| Kaiwhekea credit Cruickshank & Fordyce from Plesiosauria.com |
Ok, but what about the other trap guild specialists? Cryptoclidus is probably the best known. It had a more rounded profile, relatively larger teeth, and seems more of a generalist overall than aristonectines. It did have a wide "toilet bowl" mouth and the teeth did intermesh so it could have possibly strained food from the water or sediment but overall seems less dedicated to this lifestyle than aristonectines. My thoughts are the same for Kimmerosaurus. Kaiwhekea (above) is a strange bird. I hesitate at putting it at the level of dedication comparable to Aristonectes for this lifestyle. The teeth certainly fit the bill but what trips me up is the relatively high head. If it was feeding like Aristonectes I would expect a pretty similar low, long, and wide head. Maybe sieving infaunal prey was more its style? I have discussed Tatenectes before but dietary remains (hybodont shark) and pachyostosis in this plesiosaur (heavy, dense bones) seem to preclude it from being a true pelagic lunge feeder/filter feeder but infaunal sifting may have been important. It should be noted that the technique I described could also be applied to infaunal sifting, simply take in a large volume of sediment into gular region/oesophagus and flush through the teeth - retaining prey.

And finally, just for fun, if you have came along with me so far in aristonectines being further along than generally appreciated in the filter feeding sweepstakes let's play a little revisionist history - speculative evolutionarily speaking. If the K/T extinction event never occurred, or if aristonectines slipped through it, it is not without reason to assume that they would have further followed this lifestyle and occupied the eco-morphospace used by modern rorquals. And so we might have had 100-200 ton krill feeding aristonectines in the ocean!!

Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot
Watch me on Deviantart @NashD1. Subscribe to my youtube channel Duane Nash.


7 comments:
Interestingly enough there were aristonectines in the New Zealand region and over the years bits and pieces of very large plesiosaurs have been recorded from here.
They are of either Campanian or Maastrichtian age (Haumurian stage) and thus too late to be Pliosaurs.
This leaves either Elasmosaurids or Polycotylids and the latter are rare this late in the Cretaceous although not unknown.
So maybe a very large filter feeding Aristonectine occurred here even in the Cretaceous.
Although of course the alternative of a giant Polycotylid re-evolving the Pliosaur ecological niche would be interesting too.
LeeB.
Nice comment LeeB. If I am correct that gives us aristonectines in South America, Antarctica, New Zealand. Sure would be neat to find out they made it to the northern hemisphere and I am not surprised that we might be seeing some truly big ones discovered. I think polycotylids would very quickly re-evolve into macro-pliosaurs if it wasn't for those pesky mosasaurs!!
An Aristonectine has also been described from Angola this year, although it was too incomplete to give a name to.
Mosasaurs were evolving rapidly into lots of different niches by the Maastrichtian and probably outcompeting lots of other animals but one of the niches they don't seem to have occupied is a filter feeder; they seem to have liked chewing things up with their teeth too much.
I suspect even the juvenile mosasaurs were providing competition and predation for the young of other marine reptiles; they would be especially troublesome for other creatures if they had a poisonous bite like other Toxicoferans.
Even specialised species of genera like Globidens and Carinodens may have been more plesiomorphically general carnivores when young and only developed the specialised dentitions and diets as they aged.
LeeB.
Yes Mosasaurs were doing all sorts of neat things by then. I see a lot of similarity in their diversification as I do with Ichthyosaurs in the Triassic/early Jurassic - with the exception that mosasaurs never went toothless and, as you mentioned, filter/trap feeding.
I think what is predominantly occurring is a lineage diminishing and going extinct and mosasaurs filling in that vacant spot. Perhaps, as you mentioned, the young being plesiomorphically generalist carnivores allowed them to quickly monopolize new ecological niches and exclude others from evolving into specialized forms. I will concede that mosasaurs ability to rapidly bolt down large prey items gave them a distinct advantage in competitive food eating competitions :') Whether or not elasmosaurs would have eventually succumbed to the mosasaur hordes we will never know. Personally I think the generalized elasmosaur adaptation afforded them a lot of plasticity in where they wanted to forage (abyssal depths to estuaries/large rivers) and sharp, heterodont teeth combined with gastric mill may have allowed them to exploit tough prey items and fleshy ones too.
Any reference/publication on that Angolan Aristonectine available by chance?
Sure it is here: http://docentes.fct.unl.pt/sites/default/files/omateus/files/araujo_et_al_2015_paedomorphism-libre.pdf
LeeB.
How big would these Campanian/Maastrichtian aristonectones have been? What are the specific fossils?
How big would these Campanian/Maastrichtian aristonectones have been? What are the specific fossils?
Post a Comment