
 |
| Skiorpovenator by Duane Nash |
I told you that this was going to be a weird one, that I will be going out on that speculative branch and perhaps sawing it off. But I believe there is a sliver of a suggestion of a possibility that those bedeviled architects of weirdness the abelisaurids are even stranger than we are already allowing them. That they may in fact hail from a primarily omnivorous antecedent - with a special dietary emphasis on various gymnosperm "fruitifications" from cycads, ginkgoes, podocarps - and that even the more derived carnivorous abelisaurids carried on with a significant component of "frugivory" and were important seed dispersers and maintainers of sauropod gardens. Long story short, that abelisaurids may in fact be secondarily carnivorous.
Was Ken Ham inadvertently prescient?!?
 |
| accidentally prescient? Ken Hams Garden of Eden Carnotaurus |
What the hell you smoking Nash? How do you get a herbivore from this animal? Look at that mug?!?
Bear with me as I make the case. I don't necessarily plan to or expect to convince you… merely open your mind to the idea… maybe.
 |
| Aucasaurus credit FunkMonk CC2.0 |
Clue #1 Those Puny Vestigial Arms
Abelisaurids have arms that make T. rex look like Arnold Schwarzenegger. If you asked an abelisaur "which way to the gun show?" it could not even point you in the direction. While tyrannosaurids and other predatory theropods like carcharodontosaurids had somewhat reduced arms they still had functional arms. We might debate what if any role they played in prey capture, if they were used in ritualized combat, mating, or primarily hauled dinosaur brisket back to feeding layers, but it is very evident that predatory theropods - with the exception of abelisaurids - could do stuff with them.
 |
| Senter 2010 |
I think that point should be iterated again become I notice some conflation of the two notions of vestigial and reduced in theropods - reduced is not the same as vestigial.
Abelisaurids arms were infinitely useless in any sort of combat - although I do hold out the possibility that they were useful in dinosaurian foreplay heavy petting sessions - and most assuredly represent a genetic defect that became established in this lineage of theropod. According to this article by Brian Switek the work of Alexander Vargas (Switek cites an unreferenced paper from 2002 although I have faith in this idea as I have heard it referenced before) suggests that abelisaurids underwent a mutation in two genes that regulate the formation of the forelimb: HOXa11 and HOXd11.
This mutation, I presume (correct me if I'm wrong in the comments), is the same or similar to the genetic defect that gave diminutive, vestigial forearms to; Limusaurus; kiwis; cassowaries; Aepyornis (elephant bird); and moas. The proliferation and transmission of a genetic defect is of no real evolutionary consequence flightless birds. In the case of Limusaurus it is trending ontogentically into a herbivorous/omnivorous lifestyle and loss of a raptorial forearm grasp is of no special consequence either.
Moving from this observation it is patently obvious that predatory derived abelisaurids are very distinct ecologically from all other theropods and derived flightless avians that underwent mutations resulting in vestigial arms or wings . It is also very probable that this mutation - or ones that result in similar outcomes - arose in many lineages of theropods. But it is striking that in only one lineage of arch-predator theropod did this mutation take root - abelisaurids.
One of these things is not like the other, one of these things is something else…
With all of those diverse lineages of predatory theropods rummaging around during the 140 million years of the Mesozoic why did solely abelisaurids take on and proliferate a mutation that, it seems reasonable enough to presume, other predatory theropods were routinely exposed to?
I know evolution is not perfect and that sometimes negative mutations do get root and proliferate. But I'm still left with the stumbling block in this scenario of other theropods reducing (keep in mind reducing is different than vestigial) forearms like tyrannosaurids and even some coelurosaurs and maniraptorans, but only abelisaurids threw the arms away to the evolutionary scrap pile.
It seems logical to me to suggest that abelisaurids accepted the conditions of this genetic defect not when they were mighty titanosaur slayers but when they were piddly omnivores yet to be kings, more prone to feast on cycad fruits than sauropod entrails. If deeper in their ecological history abelisaurids were of a more omnivorous bent that could potentially explain how such a mutation could take root in animals that did not require strong raptorial forearms for their lifestyle and is more in line with the pattern of other vestigial arm reductions in theropods that have no special use for their forearms (i.e. kiwi, cassowary, moas, Limusaurus, Aepyornis). It also explains why no other predatory theropods got vestigial arms - as primarily predators this deleterious mutation would always be eliminated from the gene pool.
Clue #2: Signals of an Ecologically Adventurous Ceratosauria Being Received From the Jurassic
 |
| Limusaurus credit Levi Bernardo CC3.0 |
After the jarring sensation from the initial wtf of the idea has washed over your brain cells what should immediately come to your mind is that ceratosaurs, it has become increasingly evident, were up to some pretty weird stuff getting into the Jurassic. Limusaurus is, for me, the gift that keeps on giving. Not only does it readily encapsulate in one tidy ontogenetic sequence the transition from a predatory lifestyle to a herbivorous one complete with parallel loss of teeth, growth of beak, and arguably loss of extra-oral "lippage" but it cries out loud and clear: "We all were not scarfing on bronto guts! You should expect more of us!!"
There is every reason to suspect that, largely obscured by the passage of deep time and lack of early-middle Jurassic exposures, there was quite the adaptive radiation of ecologically diverse ceratosaurs in that period of time. Many of which were experimenting and exploring non-traditional theropod ecologies. Could the putative ancestral abelisaurid been such a dietary deviant? Other members of elaphrosaurinae should also get a nod here as they too have been suggested to be deviating from the norm in terms of lifestyle but alas no heads have been found.
Although elaphrosaurinae are currently understood to be a sister group to abelisaurids the possibility is there that somewhere deep in the pedigree of abelisaurids is perhaps an animal more prone to frequent the salad bar than the sauropod prime rib carving table.
But wait a second we already have this animal, we have Eoabelisaurus and it is just another good ol' meat chomping abelisaurid….
Or is it?
Clue # 3 Eoabelisaurus May Not Be the Meat Chomper We All Assume It to Be
When Eoabelisaurus was first revealed it was hailed as a lazarus taxon, evidence of a cryptic lineage of abelisaurids going back 40 million years earlier than the next oldest member. Personally I was always suspicious of such a long tenure of seemingly unchanged abelisaurids. Save for the shrinking forearm nothing much seems to change from Eoabelisaurus to Rugops. Of course such stasis is not beyond the pale, perhaps that is just what happened. But I have my doubts about Eoabelisaurus. I am not in doubt that it was an early abelisaurid, I am in dount that it was just another good ol' meat chomper. The reason I say this is because of some characteristics of what we have of the skull. Or what we have left of the skull.

Like any good mystery we don't have any of the tooth row from the dentary or maxillae recovered from Eoabelisaurus. What we do have are some elements from the roof and back of the skull. And why they are interesting to me is not for what they have but for what they don't have. Missing are the thickened lacrimals, postorbitals, and overall thickening and rugosity of upper skull elements so characteristic of large predatory theropods and especially every other abelisaurid yet known. In Eoabelisaurus these elements are not thickened nor is notably ornamental:
From the paper (Pol & Rauhut, 2012):
"The skull roof is not notably thickened and no cranial ornamentation is present."
It's long been a contention of mine that the thickened and gnarly skull roofs of large carnivorous theropods primarily serve as sinks for stresses and strains incurred along the tooth row. I have wrote about this idea here and there are some converging lines of evidence that point in this direction. Such thickened skull elements are not found in theropods that transition into omnivory/herbivory from carnivory. I don't take the suggestion that thickened skull roofs were primarily used for head butting competitions seriously. No studies have been conducted pointing in this direction.
Display was a secondary function I contend. The recent paper on theropod skull ornamentation (Gates et al., 2016) linked cranial ornamentation with increasing giantism in those species that had cranial adornments. What the paper failed to do was consider that biomechanical stresses may have increased the likelihood of thickened skull roofs and consummate adornments. In fact the pattern that they elucidate - cranial ornamentation linked to increased giantism - is essentially what should be predicted if these same structures served a primarily bio-mechanical function. If the function of such skull ornamentations is primarily for display we should expect such a positive feedback loop to create the largest and most elaborate osseous display features to occur in the most derived theropods. In fact the opposite seems to have occurred. The osseous displays of tyrannosaurids, abelisaurids, and carcharodontosaurids are relatively subdued compared to the earlier and more dramatic osseous display features of guys like Dilophosaurus, Crylophosaurus, Sinosaurus, and Ceratosaurus. Guanlong, an early tyrannosauroid, has much more elaborate head crests than later tyrannosauroids. Of course I think later and larger theropods largely replaced osseous display features with more dynamic, striking, and communicative soft tissue features (hello flesh antlers) but that is for another time. The paper also made the error in positing that Acrocanthosaurus lacked osseous cranial adornments & thickened skull roof when it most obviously does. In fact all large, macro-predatory ziphodont skulled theropods have thickened skull roofs often times parlayed into cranial adornments. Certain tyrannosaurids and abelisauruids even fused some of the skull roof bones together for better stress absorption. All known abelisaurids had such gnarly, thickened skull roofs. All except for Eoabelisaurus that is.
Now you may take this reasoning with a grain of salt, the jury is still out on what was the reason for thickening of the roof of theropod skulls. But what can't be disputed is that Eoabelisaurus differs fundamentally in this aspect from not only all other abelisaurids but all other large predatory theropods as well. Eoabelisaurus was perhaps not the rugged and strong biter that later abelisaurids were. It was possibly not wrestling large sauropods with that jaw. This is the best why? that I can currently come up with.
Clue # 4 Those High, Short Snouts
It has long been recognized that abelisaurids have some freakin' weird heads going on. The contrast in skulls is striking against other predatory theropods. Other large predatory theropods have narrow, long, and low snouts while abelisaurids have high, wide, and short snouts. Traditionally this difference has been explained as an adaptation on the part of abelisaurids to bite on and hold to prey as opposed to bite and slash of other theropods. My question is what is the best exaptation that would set this trend in motion?
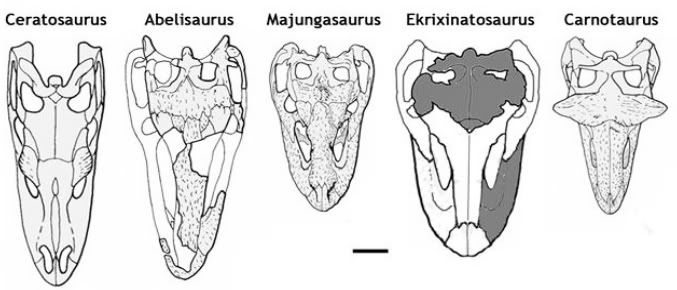
A potential explanation that may have initially shifted abelisaurids to this shape is that it was not selected for under the auspices of a hyper-carnivorous ecology but a selective omnivore. Such a short blunted skull could better pick, pluck and select choice bits of plant matter (especially reproductive propagules), small animals, eggs etc etc. Sort of like the skull of Avimimus but still toothy and retaining some predatory ability. Later on in their evolutionary tenure when abelisaurids ramped up their carnivorous inclinations the short high skull was an exaptation towards their divergent jaw and biting style among theropods. Omnivory offers a potential explanation for why abelisaurids jaws and biting style are so different from other carnivorous theropods.
 |
| credit GhedoGhedo CC3.0 |
Clue #5 Gymnosperm - Titanosaur - Abelisaurid Ecology
 |
| educational use. Podocarpus nakaii credit Ming Weng |
Of course I should stipulate that a putative omnivorous ancestral abelisaurid was always a herbivore with second thoughts. They never committed full hog to herbivory; probably did not take up gastroliths; did not digest cellulose; they obviously did not push back the pubic bone and expand the guts to ferment large piles of roughage. Their primary exploration of herbivory I suggest would have consisted in the high quality reproductive propagules, the "fruit" if you will of such gymnosperms as cycads, podocarps, and ginkgoes. Plants that were very emblematic of Gondwana and that in a diffuse coevolutionary feedback loop may have enlisted the help of abelisaurds and other tetrapods to swallow, distribute, and germinate their seeds. This coevolutionary relationship may have persisted in the younger, more derived, carnivorous abelisaurids. While Laurasia was exploding in theropod and ornithischian diversity it always seemed like Gondwana was a bit of an ecological throwback to late Jurassic times. Indeed the stasis of the gymnosperm - titanosaur - abelisaur ecological relationship that seems to have stuck around for a long time in Gondwana may in part be due to abelisaurids helping maintain sauropod gardens. Indeed where better than the rich phosphorous dung of a carnivore - even better than herbivore dung - to germinate from.
Here is a good review of the "false fruits" of the Mesozoic.
 |
| credit Brewbooks flickr. Lepidozamia peroffskyiana cycad |
This omnivorous heritage and potential maintenance would have potentially caused a receding "lip" at least on the upper jaw to allow better fine tune biting and grasping of small reproductive propagules. The lower jaw may have retained a bit more of the "meat-curtain" look, especially as large lower lips would have aided in tactile input while engaging with large prey via a neural net.
What would such a putative ancestral omnivorous abelisaurid have looked like? Probably not too different than derived abelisaurids. Actually I would take the general body plan of abelisaurids and plop on a head a little smaller, less muscular, no thick skull roof, little cranial ornaments, teeth fairly small and homodont although not necessarily non-serrated, and give it good running legs as it is now a prey animal. Such an animal was probably gorging on gymnosperm propagules when available but during the offseason dining on eggs, small game, hatchling dinosaurs often swallowed hole, carrion and just taking it easy living the easy life. We do know that abelisaurids grew pretty slow and possibly had a moderate paced mesotherm lifestyle (Ratsimbaholison, 2016). Indeed this slow paced, boom and bust lifestyle may have served these animals well. The ecology of cycads which are likewise slow metabolism plants (yes plants have metabolisms) may in fact be mirrored in one of their chief dispersal agents. Just lounging around, soaking up rays, waiting for either the next crop of cycad fruits to come into mast, the next egg laying season, or the next titanosaur to drop dead. You know, living the bear necessities.
 |
| hypothetical ancestral omnivorous abelisaur by Duane Nash |
The notion of secondarily carnivorous abelisaurids - and other "carnivorous" theropods - that augmented their mainly carnivorous diet with various high quality plant propagules is adventurous but not really beyond the pale when we put the idea in context. Ecological slosh between carnivory and frugivory is certainly a trend and frugivory seems like an ideal transitional stage to more dedicated realms of herbivory. We now know of crocodiles eating fruit and who knows how often this goes on in the wild? Specialist fruit eating monitor lizards are definitely a thing. There are several vultures that augment their diet with fruit and specialists like the palm nut vulture. Mammalian carnivorans flip flop between eating fruit and animals all the time, some becoming ultimately more tied to vegetable resources. Maned wolves are now known to be particularly fond of fruits. On the other hand polar bears are a good example of a secondary reversion to a mainly carnivorous diet from an omnivorous brown bear antecedent.
I also should at least mention the often noted rotten flesh/cheese smell put off by ginkgo fruits. The mind intuitively drifts to ideas of these Mesozoic relicts enlisting carrion glutton theropods to help consume and disperse seeds.

Rugops The Inflatable Face Monster
For having such a cool sounding name Rugops is one of the more vanilla looking abelisaurids on first impression. I mean don't get me wrong, it still had the rugged textured skull and strange proportions of abelisaurids but it was no Carnotaurus. First impressions can be deceptive however and if we peer closer at the skull roof - which compared to other abelisaurids is pretty bereft of ornaments or gnarly rocky outgrowths - there is evidence of potential for soft tissue ornamentations.

This evidence comes in the form of parallel rows of foramina on either side of the dorsal aspect of the skull. These foramina, quite apparent on the top picture, could have fed display structures. I'm not the first to notice this, Paul Sereno has speculated as such and taken note of them, although if I recall he thinks they might be keratin outgrowths. Additionally the dorsotemporal fenestra show adequate size and shelving to support additional soft tissue structures ala flesh antlers.
While many in the paleo-community (at all levels) have been generally recalcitrant to acknowledge and entertain the possibility of such features, this conservatism is not surprising to me. And you know I'm not waiting for them either or asking for their permission either. Give that Rugops a crazy looking engorged facial tissue nightmare rape face.
 |
| Rugops Tentacle Face by Duane Nash |
As I mentioned earlier when discussing the paper on osseous display features linked to theropod giantism (Gatres, 2016) why would such osseous display features - if they were so significant in spurring theropod evolution and giantism - seemingly diminish in size over the course of theropod evolution? The answer, I posit, is that such features did not so much as diminish in size but become replaced by more useful flesh and skin derived display features. Such features have the advantage over osseous display features in that they can; become engorged with blood thus changing size, shape and color and highlighting changes in mood and intent; can become damaged and re-heal, unlike osseous crests that - if bitten into - can open up the skeletal system to infection BAD!!; and serve an enhanced thermal function.

You Don't Want To Get Bit By an Abelisaur
I have to admit to developing a heavy dose of skepticism with regards to any study that gives "teh absolute bite force" of an extinct animal. I consider this akin to studies that purport to give "teh absolute speed" of an extinct animal. What I do consider such studies useful for is to give us a good range of powers/speeds but to posit an absolute number is untenable in my estimation and somewhat presumptuous. First of all let us remind ourselves that coming up with really good bite force numbers and speed estimates for living animals is notoriously hard and often changes. That should be mentioned. Secondly when we get past all the metrics, fancy math, moment arms, and finite analysis we are always left with a basic physiological question - are we talking about slow twitch muscle or fast twitch muscle? More to the point what is the relative abundance of either type of muscle - slow twitch giving more of an endurance type benefit and fast twitch more of an explosive burst of power type benefit. There is no way to decipher the relative abundance of those two muscle types from the fossil record. It is their interplay has a profound influence on questions of power, speed, endurance etc. etc.
Discussion on abelisaurid biting style are all over the place; they are called quick weak biters; sometimes Rugops (whom always seems to get picked on) is called an obligate scavenger; some call them tyrannosaurid mimics; Carnotaurus is called a weak and quick biter of only small prey. What and how were those maws operating?
I actually think that the wikipedia page on Majungasaurus summarizes the basic abelisaurid biting style quite nicely (seriously some dino-wiki pages are becoming fantastically informative kudos to the people putting in work). They were bite and hold predators. A strange paradox of the skull is featured in these animals - unlike any other predator. A thickened and stout upper jaw, a pug nosed crocodile if you will, combined with a lower jaw somewhat more slender looking and kinetic, like a snake. While struggling with prey the upper jaw could withstand extreme stresses while the lower jaw would bend and flex to prevent catastrophic damage and fracture. A rather neat and ingenious combination that differs from all other theropods and all other tetrapods that I can think of. That's my thinking as of now. Not my idea, I just happen to agree with it.
What about that small prey specialist niche for Carnotaurus? I think the idea should get some attention because the teeth in lower jaw look fairly slender, however they are not really that bad and could have been strengthened by being partially enmeshed in thick gums. More so than that the neck is just tremendously powerful looking and it has no other attributes of a small game specialist. On an ecological note where are all the modern large terrestrial small game specialist predators? Seems to me once you get much bigger than a coyote you tend to go after animals as big or even bigger than yourself - at least terrestrial predators. I mean there are no lion or bear sized mammalian predators that eschew hunting ungulates and only chase rabbits? Reminds me of the story of phorusrhacidae evolving two foot long skulls and growing eight feet high just to chase big rodents and somehow at high speeds have the skill and preternatural accuracy to conveniently slam their beak tips down on the back of the skull of their small, more agile, zig-zagging fleeing prey. Yup, I think the notion of giant terrestrial small game specialists is exactly what it looks like - a fairy tale - for both Carnotaurus and terror birds. Gigantic small game specialists are a thing in the water, but due to the unforgiving nature of moving around at 1 G on land large sized small game terrestrial specialists I would put forth as a fairy tail that, unfortunately, a lot of teh professional believe in (he-he-he). It's just not a physically and ecologically tenable existence.
 |
| credit Canale et al. 2008 |
Check out the skull of my favorite abelisaurid, the little heralded Skiorpovenator. Not only is it fairly complete, it has that gnarly bony buttress above the orbits and it has just a weird looking freakish jaw geometry. It also has what looks like some attachments for some fairly large pterygoideus muscles on the lower jaw which what I gave it in my depiction. I would not want to get bit by an abelisaurid.
 |
| Skiorpovenator by Duane Nash |
Well that about wraps it up for this one. I can't help but remember when first learned about abelisaurids as a young lad reading Gregory S. Pauls Predatory Dinosaurs of the World. So little was known about them then and it is interesting to see just how many specimens have came to light since then, and what still awaits.
The end… for now.

refs
Agnolin, F., & Chiarelli, P. (2009). The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution Paläontologische Zeitschrift, 84 (2), 293-300 DOI: 10.1007/s12542-009-0044-2
Gates, TA, Organ, C & Zanno, L. (2016) Bony cranial ornamentation linked to rapid evolution of gigantic theropod dinosaurs. Nature Communications 7. September 27, 2016. online
Pol, Diego & Rauhut, Oliver W.M. (2012) A middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs. Proc. Biol. Sciences August 2012 279(1741) 3170-3175 online
Senter, P. (2010). Vestigial skeletal structures in dinosaurs Journal of Zoology, 280 (1), 60-71 DOI: 10.1111/j.1469-7998.2009.00640.x
"A Long habit of not thinking a thing wrong, gives it a superficial appearance of being right, and raises at first a formidable outcry in defense of custom". Thomas Paine
Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot


15 comments:
You scared me at first with the notion of omnivorous Abelisaurids but it makes perfect sense as you read along. It really does explain a lot, and fits perfectly with the rest of the crazy Ceratosaurs. And I've had the same thoughts on Abelisaur biomechanics and fleshy adornments.
On another note, you bring up a possibly profound aspect of Theropod ecology and evolution: seed dispersal and the transition to herbivory. Most of the seeds and "fruits" in the Mesozoic weren't sweet and "vegan" like modern fruiting plants. They smelled and tasted far more like flesh. This could and would attract various carnivores, and therefor drive the evolution of omnivory and herbivory surprisingly and relatively quickly in the Mesozoic. That's probably why there are so many lineages and offshoots of oddly omnivorous and herbivorous Theropods and possibly omnivorous Heterodontosaurs.
One question, but where does Ceratosaurus and Genyodectes fit in with the rest of the oddball Ceratosaurs and Abelisaurs?
So abelisaurs underwent evolutionary ontogeny?
@Khalil. Yes, in may ways this is a prequel to a future post on why and how so many theropods went omnivorous/herbivorous. I think that various plants enlisted them for seed dispersal/propagation and also they were seeking carotenoids for display coloration. Stay tuned.
On Ceratosaurus and Genyodectes (quickly does a wikipedia search on Genyodectes as i totally forgot about that guy). I don't really have anything to say on those guys new. I'm not sure what your question is but, as I said in the post there is every possibility that there was a large and diverse radiation of ceratosaurs in the Jurassic. In many ways this radiation might parallel and presage the large scale radiation and diversification of maniraptorans which we have better evidence for due to more recent and widespread fossil deposits. Early and mid Jurassic ceratosaurs certainly hold a lot of promise for unexpected surprises.
@ Bk Jeong Not sure what you mean… is that with regards to head shape?
Fascinating thoughts here.Have you seen Mark Witton's latest post? It also concerns *Rugops* and deals with soft tissue ornamentation and dermal armour in abelisaurs in general.
A minor point here, though: moa did not have vestigial forelimbs; they lacked forelimbs and even a pectoral girdle altogether.
Your (originally) omnivorous abelisaurs remind me of the fact that most large, flightless birds went for big heads and beaks. It's a pity most surviving examples are ratites as these are atypical in being pinheads. Takahe (and related, extinct types), kakapo, adzebills, phorusrhacids, dromornithids, sylviornithids, gastornithids, dodos, *Cnemiornis* and moa nalo all come to mind. As such, developing or retaining large noggins seems a pretty strong trend among theropods that evolve into large, heavy, terrestrial omnivores/herbivores. (Though admittedly, omnivorous/herbivorous coelurosaurs offer counterexamples i.e. ornithomimids and therizinosaurids) Abelisaurs may be another and very ancient example. Though the resemblance is probably superficial, I find large dromornithids to have beaks that particularly look like abelisaur snouts in that they are proportionally big, high, domed and mostly broad but narrow at the tip.
Where do you say noasaurids fit into the picture? They are, at least sometimes, regarded as being closer to abelisaurids than to *Limusaurus*,*Elaphrosaurus*, *Deltadromeus* or *Ceratosaurus*, right?
In general, the main predatory organ in theropods is the skull, so I see no particular problems in abelisauroids losing forelimb function at all. They used the neck-mandible system in predation, as all other carnivorous dinosaurs. Note that abelisauroids (both abelisaurids and noasaurids) show a very complex pattern of epiaxial muscular sites in the neck, which suggests a neck musculature more developed than in other theropods: this compensated forelimb reduction. The stronger head-neck complex acted as forelimb in prey capture and manipulation, with no need for appendicular help.
Eoabelisaurus: there is intriguing evidence that it may be a ceratosaurid, thus a genuine predator (pending direct evidence from dentition). The skull elements known are unornamented and with thickness comparable to Ceratosaurus: so, if we assume the latter was a predator, why not Eoabelisaurus too?
The nasal foramina in Rugops are reported also in Carnotaurus. It is unclear if they were pneumatic (and thus may support inflatable structures) or just neurosensory. Given their size and regular pattern, I bet they are neurovascular and not pneumatic (that usually show a more random pattern and larger size).
There is no particular problem in widespread omnivory among theropods: whatever could be ingested whole and does not require chewing represented a potential food item for these animals. So, I am skeptical on this ad hoc speculation for abelisaurids.
@BrianL Yes I saw Mark's post. It was an interesting coincidence but I have had similar convergences many times while blogging. Interesting observations about head size. Will keep that in mind.
@Andrea Cau Indeed skepticism was to be expected. My main point with the forelimbs in abelisaurids is that they were not merely reduced - they were vestigial. There is a difference. Why, how, and when the interesting neck and jaw adaptations arose in abelisaurids is a different question than why was a deleterious mutation allowed to proliferate in these animals. Many other theropods show reduced, but not vestigial, forelimbs. Why did this mutation take root in only abelisaurids? It could be a genetic quirck, some sort of genetic bottleneck a foundational population took on. Or it could be that more basal abelisaurids were of a more omnivorous inclination and raptorial forelimbs were not important (as seems to be the case for Limusaurus). Is my explanation ad hoc? Of course it is. But is the other explanation, a genetic bottleneck, also ad hoc? It is as well.
Interesting about the nasal foramina on Carnotaurus. If they were neurovascular I wonder their purpose, seems a bit of an odd spot? Also interesting about Eoabelisaurus as a ceratosaurid.
"There is no particular problem in widespread omnivory among theropods" Agree there.
@BrialL "What about noasaurids?" Great point, I actually should have discussed them in the post. Masiakasaurus has fully functional forelimbs in fact they may have been useful at expanding and digging into lairs of fossorial prey. So we can't just say that you lose your forelimbs because you are in the clade abelisauria. What is more likely going on IMO is that you lose your forelimbs because you don't have a use for them. Abelisaurids remain the only theropods (ground birds included) that got vestigial forelimbs and were predatory. Other predatory theropods reduced forelimbs, but they did not go vestigial.
That is an excellent point on vestigial versus reduced. The function of theropod forelimbs has really intrigued me lately. Despite a very strong selective pressure to reduce the forelimbs for biomechanical reasons, there nevertheless seems to be an equally strong ecological pressure to wring every last possible bit of use out of them. Hence even tyrannosaur limbs are pretty beefy for their size.
However, despite being the one clade with truly vestigial limbs, even the abelisaurs seem to still be getting SOME use out of them. Just look at the size of their shoulder blades! I don't know much about biomechanics, but supposedly they have unusually mobile shoulder joints as well.
I meant that they reverted to being predatory
@Andrew Raymond Stuck on vestigial versus reduced. The concept that reduced is not the same as vestigial is not mine. One of the researchers who looked at forelimbs in theropods/birds iterated it. The distinction bears repeating however, as too often reduced is conflated with vestigial. On abelisaur shoulder blades. I know!! they are like crazy robust. I don't know what to make of them, the use I often see mentioned and which makes sense is the they anchored the heavy neck musculature.
This whole discussion of predatory theropods forelimbs has definitely sparked some new thought in my mind as to what other large theropods were actually using their forelimbs for. I will offer this up in a future post(s)? But I think giving the obvious problems that arise in theropod forelimbs it will prove useful to attach the issue from a different angle.
I wrote about vestigial vs reduced arms in theropods a lot of years ago:
http://theropoda.blogspot.it/2008/10/perch-il-braccio-di-tyrannosaurus-non.html
All non-abelisaurid short-armed theropods had functional forelimbs involved in predation.
Even tyrannosaurus forelimb is used in predation, more or less the same way as other tetanurans used their forearm in predation: as a tool that blocks the prey to help the head-neck system to deliver a series of bites. The weaker the bite (and so, the more numerous the bites necessary for killing), the stronger the forelimb necessary to block the prey. The stonger the bite, the weaker the forelimb because it was not necessary to block much the body of the prey killed by a few bites.
Following this pattern, abelisauroids are easily explained:
Abelisauroids lost forelimb use and further increased neck musculature, because they changed predatory style maximizing bite impact: in fact they evolved short snouts, larger infratemporal and mandibular fenestrae compared to other theropods, thick skull bones, short and robust teeth, and developed the epiaxial neck muscles. This is an adaptation for a small prey that is killed with a single fast bite delivered by a quick head moved by a strong neck. No need to use the forelimb as instead happens, for example, among tyrannosaurids that need the forelimb to assist the bites over a large prey.
If forelimb atrophy is linked to omnivorous-herbivorous habits, where are the vestigial forelimbs among ornithischians? Where are among omnivorous maniraptorans? The latter show exactly the opposite.
Also, the hand of Ceratosaurus is still functional, based on metacarpal facets. The hands of noasaurids retain claw-like unguals. The hand of Limusaurus has been shown to be a parallel reduction, not the ancestral abelisauroid condition. Only abelisaurids show the full atrophy, and it is interesting they are those that evolved the short and muscular neck, the short and thickened skull and the robust dentition.
@Andrea thinking about theropods arms has caused me to seriously reconsider their role in predation. I think that their arms had a use, but it wasn't primarily prey capture. I will revisit this idea in a future post and also discuss and probably think more about your comments on ornithischians/maniraptorans etc etc. Thanks for your thoughts!!
I don't think theropod arms were used in prey capture: the mandible was the organ for prey capture. The arms helped in keeping the prey after the mandible had captured it.
This explains why in predatory theropods the medial most finger (pollex) is always the strongest and is oriented medially.
Just a note: as a paleontologist, I welcome any speculation. But to be a scientific hypothesis, such speculation needs to be tastable. For example, a functional hypothesis should be consistent with a phylogenetic pattern built independently from the scenario suggested. Your erbivorous/omnivorous idea for ancestral abelisauroids is not supported by any current ceratosaurian topology.
I have to side with Andrea on this one. Severely reduced forelimbs have evolved many times within Theropoda, in ecologically disparate lineages: insectivores (alvarezsaurs, flightless rails), herbivores (elaphrosaurs, moas, elephant birds, flightless waterfowl), omnivores (basal troodonts, some microraptorids), and obligate carnivores (ceratosaurs, abelisaurs, carcharodontosaurs, tyrannosaurs, advanced troodonts, hesperornithids, gastornithids, phorusrhacids). The functional reasons for forelimb reduction are not always clear cut, and highly doubt that in the case of ceratosaurians, it had anything to do with omniherbivorous habits (after all, baby elaphrosaurs were sharp-toothed and probably carnivorous, unlike the herbivorous adults, and they too had reduced forearms).
While I was reading this, I kept thinking of a sentence from the abstract of Gould & Lewontin (1977): "Male tyrannosaurs may have used their diminutive front legs to titillate female partners, but this will not explain why they got so small." Adaptationist thinking has its limits.
All that being said, I do like the idea that stereotypically carnivorous theropods may have occasionally consumed vegetation, like modern carnivorous mammals do in order to ameliorate health problems (i.e. dogs eating grass). And I'm glad you dispel the myth of abelisaurs as weak biters (I'm especially sick of Rugops being portrayed as an obligate scavenger); they must have been powerful enough to attack the huge sauropods and fast-running thescelosaurs, and successfully replace carcharodontosaurs as Gondwanan tyrannosaur-analogues.
Post a Comment