
Recently I went out for a hike in the local Santa Monica mountains. Spurred on by the recent heavy winter rains in California I was expecting a bouquet of abundant wild-flowers. The flowers were there of course and it was breathtaking. However as the strange has an unusual affinity for me (and I for it) what got my attention as soon as I opened my truck door at La Jolla canyon was not the flowers, but a dead hatchling theropod on the ground. Not sure what species - probably a larger corvid due to size - but as I peered closer I noted a strange bundle of flesh at the crease of the mouth.

Some genetic aberration? The avian version of a cleft palate? Or perhaps a link to a more primal ancient pedigree of lipped theropodian monster faces? There is a saying in evolutionary theory: ontogeny recapitulates phylogeny. Is this a case of that? I looked at lots of pics of hatchling birds online and can't find any with that degree of bunched up oral tissue at the commissure. Whatever your thoughts on what this might represent - probably is some genetic defect - you have to admit the weirdness of it all… a dead beaked bird with a fleshy extension of the oral margin directly reminiscent of the very same depictions I have been advocating.
The picture below is a screen shot from a youtube post by C.M. Kosemen detailing his thoughts on the chickenosaurus. The pic is actually taken from a book titled Incredible Life: A Handbook of Biological Mysteries by William Corliss, purportedly a rare treasure trove of arcane biological curiosities described by Kosemen as " the Necronomicom of biological weirdness". According to Kosemen the book documents a chicken from 1878 born without a beak and having strange "mammalian like head features". The oral tissue surrounding the mouth described as a quasi gummy but tough tissue.

Thanks for commenter Casielles for bringing this pic to my attention and noting how much it resembles some of my lippy theropod designs.
Wherever you fall on the chickenosaurus debate, the Dr. Moreau in me is still curious about what sort of oral tissue would come about on a chicken reverse engineered and stripped of its more derived facial attributes into something well… not quite a dinosaur... but not a typical beaked bird either. Could we garner a glimpse at the primordial oral margin of theropods prior to the evolution of beaks?
The photo below is credited to Bhart-Anjan Bhullar and shows how through suppression of certain proteins the premaxillary bone of a chicken (labelled experimental) can be suppressed giving arise to a more basal configuration.

While the fusion and expansion of the two premaxillary bones into the "beak" is certainly interesting, all of this does beg the question: if beaks can be reverse engineered into snouts can we also reverse engineer the oral tissue that lines the mouth of birds into what once was?
Bhullar et al. (2015) isolated specific proteins that stimulate the development of beaks (i.e. the premaxillary bone) in modern birds. Although I can find no pictures of the creature that they came up with - from what I gather the birds were not allowed to hatch - they were reportedly able to control these proteins and in doing so reportedly come up with a chicken closer to the ancestral state in terms of facial bones and lack of beak. What they said - and what caught my interest - is that this manipulated chicken skull looked superficially normal from the outside but something obscured a clear look at where the beak should have been. What was obscuring the beak area, you ask? A flap of skin was. Was this flap of skin a reversion to a more primordial "lip" condition? I don't know. What is interesting in this study is that another unexpected osteological change occurred - the palatine bones reverted to a more ancestral condition. Manipulation of one aspect - the beak - can have unexpected correlates in another osteological condition and I'm asking maybe even supporting soft tissue components as well…
 |
| credit Bhullar. Showcases loss of beak creates ancestral palatine condition |
Take what you will from these studies, they of course have been met with loads of skepticism. But I do have to wonder if they hint at a pathway to a closer approximation of soft tissue oral margins in theropods.
Croc Faced Killers
 |
| Croc-Faced Lungfish Fighting Mud Frolicking Spinosaurus by Duane Nash |
Of course for those keeping abreast of things it is all about crocodile faced theropods right now. Yep, rugged, tooth exposed, thin lipped, croc faced killers. And let's give credit where credit is due: Tracy L. Ford has been sounding off on this look for years before the Carr paper hit. You know some might think I am against the notion of croc faced theropods - quite the contrary actually. I do want to go on record saying that the croc faced idea and lack of lips notion in theropods is a good one and a very important concept and likely prevalent in a great many theropods - just not for all of the theropods that this look is normally attributed to.
A necessary prequel to the ideas I will be elaborating on is my last post Behind Your Bony Mask of Face. In this post I made the argument that the texture of the skull of crocodiles has more to do with ecology than it does skin integument, that in fact a similar skull texture is found on many amphibious and aquatic stealth predators - some that have tightly adhering skin and some that do not. What tyrannosaurids like Daspletosaurus horneri and indeed most carnivorous theropods have is indeed a rough and textured skull that, I suggested, helps anchor a particularly tough and textured epidermis. This dermis - possibly even somewhat keratinized and/or cornfield - is not necessarily covered in scales.
Is there a test of this notion of mine? Can we locate an animal - preferably somewhat close ecologically, behaviorally and phylogenetically to tyrannosaurids - that shares such a rugose and textured skull BUT that does not have a scaled facial epidermis?
By golly, yes there is and it comes in the form of a lineage of theropods that are extinct, were arch hunter-scavengers, did have nasty biting abilities, most certainly face bit, were not aquatic, and were most certainly not scaled in the facial region. The answer is phorusrhachoid terror birds.

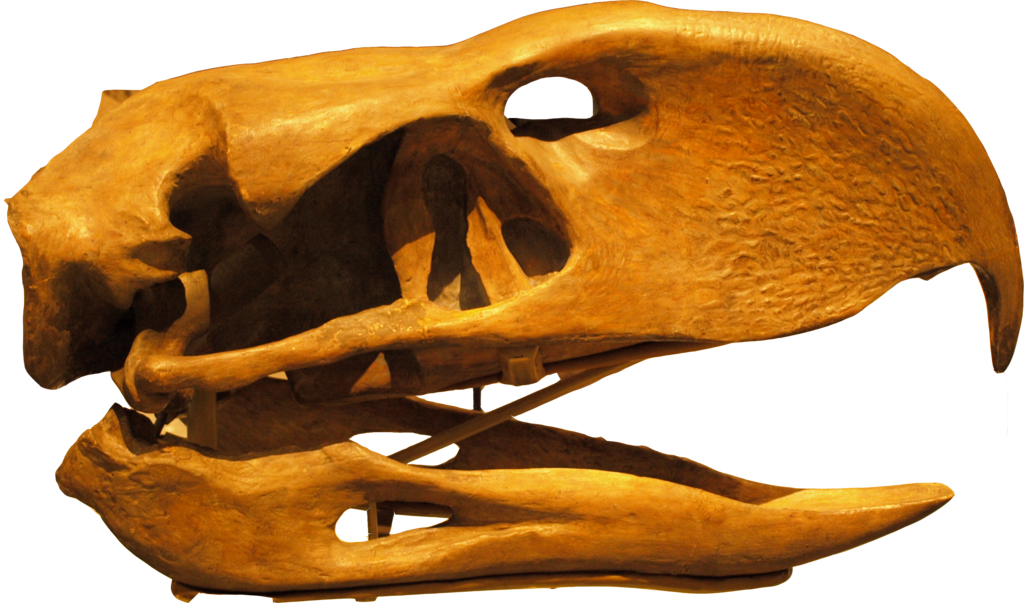 |
| Phorusrhacos longissimus Royal Ontario Museum, credit Captmondo CC3.0 |
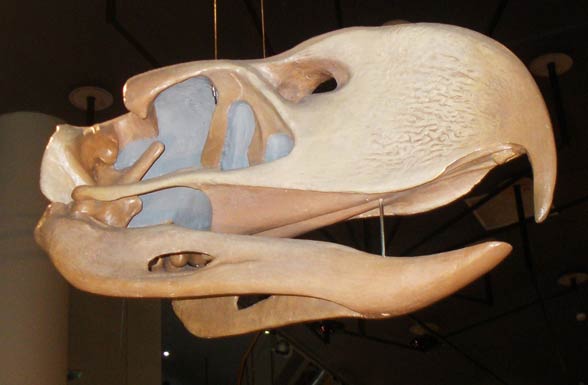 |
| Phorusrhacos from here |
 |
| AMNH 5027 public domain |
What I am suggesting is that the facial osteological correlate of these animals is not so much a phylogenetic signal, but an ecological one. Large carnivorous derived theropods (i.e. phorusrhacoids) having a textured skull convergent with their ecological antecedents in large carnivorous extinct theropods. A necessary caveat of this is that, as I mentioned, those theropods that most converged closely with crocodilians ecologically likely had a croc - like oral margin and skin texture. These theropods would most notably include spinosaurids. Other small game theropods especially of the "kink - snouted" gestalt may have been similar or split the difference: lack of extra-oral tissue towards the front of the jaw but retaining some "lip" towards the rear of the jaw. Noasaurids which were always scrounging into the dirt after fossorial prey may have diminished lips as well. Many smallish coelurosaurs, dromies, troodontids, and compsognathids that were small prey scroungers may have followed this pattern of diminishing, receding lips and a more croc-like exposed oral countenance.
Toothed Birds, Pseudo-toothed Birds and Small Game Foraging To Lose Lips
 |
| Sinosaurus credit Duane Nash |
 |
| Sinosaurus credit Ghedoghedo |
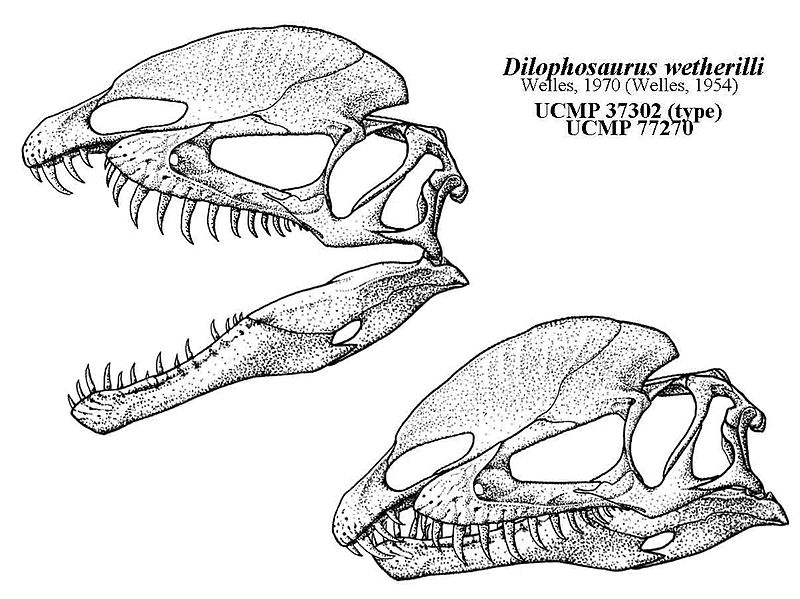 |
| - http://qilong.deviantart.com/art/The-Dilophosaurus-Yawn-252396964 |
Again with Dilophosaurus it is simple to formulate an ecological reason for it dispensing with lips. Because it shows tendencies towards small game, large lips in the anterior of the jaw would provide a cumbersome block to intricate poking and plucking of small critters whether in the water, in burrows, or whatever. Better to dispense with such lips, evolve a keratinized and/or cornified oral epidermis epidermis and retain sensitivity from the neurovascular foramina. Note how this parallels the argument in my last piece in this series where I suggested that the evolution of beaks and loss of teeth were coincident with an increasing emphasis of omnivorous/herbivorous diets. Where as the constant pecking, plucking, and cropping of seeds, fruitifications, foliage, insects would encourage a beak type oral margin to replace non-muscular fleshy "meat curtain" type oral margins in theropods: in croc-faced killer theropods the lips would likely recede for the same basic reasons. In these small game hunting theropods the constant abrasion against substrate, the drag imposed on lips in water, the delicate plucking of small game would encourage a loss of extra-oral tissue. Like theropods that evolved beaks, theropods that moved towards specializing in small game, burrowing animals, "piscivory" would lose lips but they would not lose their teeth.
A constant argument I put forth for large lippy extra-oral tissue in large game theropods is that this tissue provides an extensive "neural net" that allows real time sensing of the movement of large prey in the jaw and corresponding bite adjustments. For theropods foraging for smaller prey in the water such a "neural net" is not needed. Water itself provides the medium for sensing prey movement and as the lips would hinder small game plucking, encourage drag, and are altogether redundant they would soon diminish and take on a "croc-like" oral gestalt. The sensitivity remains however, it is just the lips that recede.

It is also possible a very tough gummy type tissue covered up a lot of the tooth in Dilophosaurus. Such gummy tissue would of course be a lot more rugose and resistant to drying than typical gingiva but still anchor and strengthen the teeth.
Teeth, or better yet pseudo-teeth are important for both extinct and extant proper aves and near birds like the often toothed enantiornithines. There are many examples of the evolutionary utility of teeth, choannal papillae, and pseudo teeth in these animals showing us that not only are such projections useful and whenever possible retained BUT more importantly, I would argue, that there is no universal a piori directive, guidance, or eventuality of an edentulous "beak" in theropods. These examples also fundamentally address the importance of ecological consideration when thinking about extra -
oral margins and "toothiness".
 |
| Pelagonis mauretanicus crédit Didier Descouens CC4.0 |
 |
| Pseudo teeth penguin credit: http://i.imgur.com/o3vze.jpg |
To bolster this pattern simply look towards the dominant fish eating birds around in the Cretaceous: hespernornithiformes and ichthyornids both have toothed jaws. They had basically lipless keratinized beaks - as should occur given their predilection for aquatic prey. Yet they retained teeth. And there is no reason to suspect that they would lose their teeth if the K/T event did not occur.
 |
| Hespernornis regalis YPM 1206 credit Heilmann 1926 |
It is worth mentioning that in the above species it has partially lost teeth in the upper jaw, replaced by sockets that the lower teeth slot into. Ichthyornis shows a similar pattern: tooth loss partially in the upper jaw but a continuous row of teeth in the lower jaw. One has to wonder if and why we should expect complete tooth loss in these groups given the propensity for truly toothless birds to evolve pseudo-teeth.
 |
| Ichthyornis public domain credit O.C. Marsh |
 |
| credit Stephanie Abramowizc |
 |
| Sulcavis geeorum credit Stephanie Abromowicz |
Jingmai O'Connor:
"While other birds were losing their teeth, enantiornithines were evolving new morphologies and dental specializations. We still don't know why enantiornithines were so successful in the Cretaceous but then died out - maybe differences in diet played a part."
So where does this leave us? Admittedly this piece is a little all over the place, and readers might be dumbfounded in how quickly I switch from say a critter like Sinosaurus to Pelagornithis. But there are some general themes and trends that stand out.
- The transition of toothed birds to toothless bird is not a direct line nor is it necessarily inevitable. The prevalence of pseudo-teeth in many modern birds (especially piscivorous) that correspond to toothed Cretaceous analogs should give us moment to pause. Perhaps the Cretaceous toothed ichthyornids and hesperornithiformes with their basal retention of teeth were not the inferior models compared to modern penguins and "gull" type seabirds but actually the optimal model due to their teeth which should outperform the pseudo-teeth of modern birds.
- Enantiornithines provide us with an alternative universe where toothy birds are not only common but the most successful "norm". We might be partially conditioned to think of modern derived toothless birds as the optimal model for what a bird should be, but what if this is not the case? Enantiormithines seemed to dominate toothless birds in the Cretaceous. Did the K/T boundary provide the ecological filter that allowed toothless seed eating birds to get through but prohibiting the more toothed - and carnivorous - enantiornithines from getting through as has been suggested in a recent paper?
- In both the toothed fish eating birds and the carnivorous enantiornithines I would put forth a condition of highly reduced lips - perhaps just a trailing edge of tissue at the commissure. This is also similar - albeit without a beak - to the near lipless condition I would expect in small game hunting, fossorial foraging, and piscivorous theropods. The realities of aquatic foraging, constant abrasion, and intricate plucking of small game would create a less than ideal situation for any substantial, non-muscular extra-oral tissue in these animals. This provides an alternative pathway to losing lips to what I discussed in my Lips Before Beaks part I where herbivory would diminish lips and create beaks in theropods.
In all the above examples I am actually explicitly arguing for an exposed toothed, primarily lipless, and even croc-like oral margin. Wait a second here… wasn't I the guy who argued for lippy sabertooth cats, beneficial activity of saliva on tooth health and what not? Have I done an about face?
Let's just say that my thoughts have… evolved and become a bit more nuanced. Two things can be simultaneously true and seemingly at odds but still coexist. Sheathed teeth under the auspices of calcium and phosphorous buffered salivary broth I still believe is a great thing to have… just not mandatory in all cases. Pterosaurs, crocs, plesiosaurs they all break that "rule" and a good many theropods probably did too… just not all of them. I would still consider most largish big game, serrated toothed macro predatory theropods to maintain - and even re-evolve - large "meat curtain" type lips. This would include most largish tetanuran type theropods, allos, carchs, predatory ceratosaurus, tyrannosauroids, dromies, megaraptors - generally most serrated toothed zyphodont theropods.
You will note I left out abelisaurids, I think something neat was going on with them which I will save for another post. Yep, some have been asking for it and I finally have enough new ideas to make a post on abelisaurids. Trust me, they will be weird AF when I'm done with them. Weirder than they already are.
And finally I leave you with Austroraptor. It was a South American unenlagine potentially flightless spinosaurid mimic. Remember spinosaurids themselves are crocodile mimics and share with them a skull texture (spinosaur skull texture is more similar to crocs than tyrannosaur skull texture is to crocs imo) and likely a lipless oral margin. Guess what the skull texture of Austroraptor looks a bit like?


 |
| Austroraptor cabezii by Duane Nash |
Austroraptor is that unique beast that potentially tells us a lot. Because it was a maniraptoran and quite possibly secondarily flightless it may indeed hail from fully feathered ancestors. If it lost feathers on the face it may have indeed followed the trend of modern naked faced birds and eschewed feathers and scales on the face. After converging with spinosaurids (which converged with crocodiles) it evolved a highly, rugose, textured, and pitted skull. Not because it had a scaly face but because such a skull texture allows - as I suggested in Behind Your Bony Mask of Face - better tactile and neurosensory capability as a sensitive snouted aquatic predator. Hence the convergence of skull features as I highlighted in such disparate groups as temnospondyl metoposaurs, phytosaurs, snapping turtles, crocodilians, various fish, and others. These textured skulls in aquatic predators I hypothesize allow more surface area to "catch" vibrational frequency in the water medium like a gnarled, old used catchers mitt.


Refs
Bhullar, B.-A. S., Morris, Z. S., Sefton, E. M., Tok, A., Tokita, M., Namkoong, B., Camacho, J., Burnham, D. A. and Abzhanov, A. (2015), A molecular mechanism for the origin of a key evolutionary innovation, the bird beak and palate, revealed by an integrative approach to major transitions in vertebrate history. Evolution, 69: 1665–1677. doi:10.1111/evo.12684
Carr, T. D. et al. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Sci. Rep. 7, 44942; doi: 10.1038/srep44942 (2017).online
Vertebrate Paleontology, SVP 75th annual meeting, Meeting Program & Abstracts,
p. 125.
O’Connor, J., Zhang, Y., Chiappe, L., Meng, Q., Quanguo, L., Di, L. 2013. A new enantiornithine from the Yixian Formation with the first recognized avian enamel specialization. Journal of Vertebrate Paleontology. 33, 1: 1-12
Weeks O, Bhullar B-AS, Abzhanov A. 2013. Molecular characterization of dentadevelopment in a toothed archosaur, the American alligator Alligator mississippiensis. Evolution and Development 15(6): 393-405.
"A Long habit of not thinking a thing wrong, gives it a superficial appearance of being right, and raises at first a formidable outcry in defense of custom". Thomas Paine
Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot
Watch me on Deviantart @NashD1. Subscribe to my youtube channel Duane Nash.
16 comments:
Well, well, that's really interesting, and opens so many possibilities when it comes to restoring theropods & friends (Austroraptor, the croc-bird?).
Didin't know about that weird 19th century chicken, do you think it could lend some credibility to the idea i had for some time, that some theropods (mainly tyrants and allos, basically the big ones) could have had some sort of mammal-like lips which could (more or less) move?
Hi Alesssio. I know that Bakker toyed with the idea of weak facial lip muscles in theropods that allowed a gentle sneer look. I have to admit to not being sold on the notion. I would like to see some exaptation that would create such conditions to arise in theropods - mammals have to suckle after all so their prerequisite for facial muscles is there - but I can't fathom why theropods would evolve muscular lips/faces when their modern relatives are impoverished in such features.
Very good and thought provoking post! I like where all these lippy ideas are going. The need to gain and lose lips due to foraging habits makes a lot of sense, and I have always wondered the oddness of the weird kink in many small to medium sized Theropods. The need to pluck prey accurately makes a lot of sense.
Many thanks for the answer, Duane. Well, about the possibility of moving lips in theropods maybe you're right, i didn't remember that we mammals have this feature because, well, we suck (man, what a lame pun!)... The only way a dinosaur could have employed mobile lips, mmh, maybe for communication. But, as far as we know, reptiles and birds have an helluva lot of ways for communicating, without making faces, so... By the way, i also didn't know it was Bakker who talked about this possibility.
where do ornithischians (cheeks) fit in with this? There's been some disagreement about what used to be the status quo since the 70s/80s (starting I think with Witmer)
@Tom Y. Good question!! Not sure myself!!
Awesome post, interesting to see your ideas shift and creating even weirder hybrid ideas, love the Sinosaurus, absolutely terrifying
On the topic of beaked pterosaurs, what kind of mouth do you think Azhdarchids had? Crazy gripping papillae, or something more like a stork, with a more or less featureless interior mouth. A while back I did a pteranodon with penguin and seagull inspired papillae http://trilobitecannibal.deviantart.com/art/Pteranodon-the-artquel-665751427
Hey Trilobite Cannibal. If I were to hazard a guess I suspect that the trends we see in most comparable analogues might offer useful clues. It should be stated that I know of no significant study linking osteological correlates to various papillae structures. this line of inquiry might offer some useful insights into the potential for papillae type structures in various edentulous pterosaurs. But, yes, I would speculate that azhdarchids likely had stork like mouths and the piscivorous pterodontids >may< have had the potential for papillae.
Dear Mr. Nash
I do like your idea of "meat curtains" but your claim that lizard style lips would not work with naked skin, as they would get in the way of biting. "Incidentally I do think lips getting cut up is a big problem in the "tooth pocket" design most notably implemented for T. rex in the game Saurian. What is preventing such a loose, non scaled lower lip from flopping inwards during the bite and getting penetrated? Scales might offer more structural support but if the face was not scaled?" But, scaleless, naked, lizard style lips exist. Scaleless pythons exist in the pet trade http://thereptilereport.com/scaleless-ball-python-first-full-pic-leaked/
And, toothed whales. Their teeth are covered rather minimally. And of course aren't scaly. http://www.mammalogy.org/orcinus-orca-1058
Both are quite capable of biting without the lip flopping inwards during the bite and getting penetrated. Am I saying your idea is a bad idea? No, I rather like it. But I am saying that scaleless naked lizard style lips do exist, and thus are still a plausible option for dinosaurs. Thank you.
@ MrCrow. good points I will look at these. I have been thinking a lot about the structural integrity of things like hippo tooth pockets which house large teeth but don't flop around.
New paper suggesting Integumentary Sensory Organs on Neovenator. And more evidence than merely skull texture this time. Suggests that the mouth covering was thick and immobile.
Neovenator ISO paper here (https://www.nature.com/articles/s41598-017-03671-3)
Thank you I have read it. Darren teased on his podcast that they had found evidence for lips in their study but that was left out in the study for some reason. I think I know what the evidence is but don't want to spoil it!!
You refer to Austroraptor as "potentially flightless". Can you elaborate on this? It was huge and had diminutive arms. Unless juveniles have been found with a totally different anatomy I just can´t imagine how Austroraptor could be anything but obviously, most definitely flightless?
Great read btw as always.
Sorry that is a bit of a confusing phrase. I am not implying it was potentially flight capable merely that its ancestors may have been flighted.
The theropod sneer? Well, at least one snake is capable of curling it's upper lip into a snarl (https://www.researchgate.net/profile/Jose_Pedro_S_Do_Amaral/publication/249488146_Lip-curling_in_redbelly_snakes_Storeria_occipitomaculata_functional_morphology_and_ecological_significance/links/590b5003aca272f6580eb5b2/Lip-curling-in-redbelly-snakes-Storeria-occipitomaculata-functional-morphology-and-ecological-significance.pdf). Then again snake skulls are very different from Dinosaurian skulls.
Post a Comment